Human auditory cortex representerer 8% av overflaten av hjernebarken. I motsetning til andre hjerneområder er det viktige strukturelle forskjeller mellom hørebarkene til forskjellige pattedyrarter, så vel som mellom aper og mennesker.
Anatomi av menneskelig auditiv cortex
Den menneskelige hørselsbarken kan studeres ved hjelp av funksjonell magnetisk resonansavbildning (fMRI), og er delt inn i mer enn et dusin forskjellige regioner som omgir Heschls gyrus i den øvre delen av den temporale loben. smalere hos mennesker enn hos andre pattedyr, utvikler seg fra front til bak i Sylvian-sprekken på det punktet der den blir med Heschls gyrus.
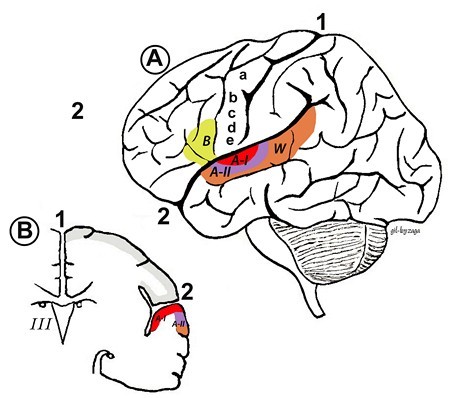
Primær auditiv cortex (AI) ligger i den bakre tredjedelen av overlegen temporal gyrus (også kjent som Brodmann-område 41), ved siden av Wernicke-området (W). AI er den sentrale regionen i hørselsbarken og mottar direkte projeksjoner fra den stigende hørselsveien, spesielt den ventrale regionen i den mediale genikulatkroppen ( MGB) i thalamus.
Sekundær auditiv cortex (AII) ligger mer rostralt i temporal lobe og inneholder Brodmann-område 42.
Anatomisk fordeling av auditiv cortex
Struktur og kretsløp i auditiv cortex: Søyleorganisasjon
Tilstedeværelsen av seks celler lag i hørselsbarken er vanlig for alle pattedyr, men artsforskjeller har form av fellesheten til hver cellene i hvert lag. Hos mennesker tilsvarer pyramideceller (inkludert alle typer) 85% av AI. De resterende 15% er multipolare eller stjerneceller. Inverterte stjerneceller eksisterer også (Martinotti-celler) så vel som celler med kandelaberformede dendritiske konfigurasjoner.
De fleste stigende fibre har opprinnelse i MGB og synaps med pyramidecellene i lag IV, men dette er ikke alltid sak. Imidlertid representerer disse kontaktene bare 20% av de eksiterende fibrene som rager ut til kortikale nevroner: de andre 80% kommer fra andre nevroner i den ipsilaterale cortex.
Nevroner i AI og AII er funksjonelt organisert i kolonner, først beskrevet av Lorent de Nó. Kortikale kolonner mottar innspill fra begge MGB og er derfor bilaterale, og arbeider med prinsippet om summering / undertrykkelse. Summasjon tilsvarer lignende afferens fra begge ører, med en kontralateral dominans. Undertrykkelse er ipsilateralt dominerende.
Mobil organisering og kretsløp i den menneskelige hørselsbarken
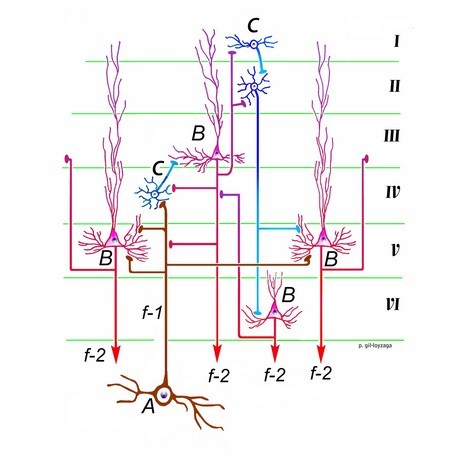
Hver neuron i MGB som projiserer til auditiv cortex (C) genererer en fiber (f-1) som forgrener seg horisontalt for noen få millimeter og kommer i kontakt med mange pyramideceller (B) og puncta (C). Dette systemet tillater forsterkning av det auditive signalet og forbedret analyse av dets aktivitet. Nevroner i lag IV rager ut til de pyramidale nevronene i lag III, og derfra distribueres informasjonen til de andre lagene (I, II, IV og V) i den ipsilaterale cortexen og den kontralaterale auditive cortexen via corpus callosum. Lag I nevroner projiserer til lag II, som igjen forbinder med lag V og VI. Pyramidale nevroner i lag V og VI har efferente aksoner (f-2) som projiserer til MGB. De i lag V projiserer også til den underordnede kollikulus. Alle disse nevronene sender også sikkerhetsforbindelser tilbake til lag III og IV.
Spesifisitet av den menneskelige auditive cortex
Mens andre nivåer av hørselsveiene er veldig like innen arter, er det humane neokortekset preget av overvekt av pyramideceller (85% av kortikale nevroner), og noen veldig spesifikke typer celler som inverterte pyramideceller og lysekroner. En annen spesifisitet er den massive sammenkoblingen av kortikale nevroner, som utgjør 80% av eksitatoriske synapser i neocortex. Bare 20% kommer fra legemiddelkroppen!
